
|
|
|
Human Nature
- Introduction
- What is Human Nature?
- Psych 101 Mind-Body
- Psych 101 Personality
- Psych 101 Motivation
- Brain Development
- Brain Learning
- Psychology Intro
- Psychology Primer
- Psychology Design
- Human Abilities
Other Pages
Evolutionary Psychology - Design of Organisms
Evolutionary Psychology: A Primer (continued from page one)
by Leda Cosmides & John Tooby
Understanding the Design of Organisms: Adaptationist Logic and Evolutionary Psychology
Phylogenetic versus adaptationist explanations. The goal of Darwin's theory was to explain phenotypic design: Why do the beaks of finchs differ from one species to the next? Why do animals expend energy attracting mates that could be spent on survival? Why are human facial expressions of emotion similar to those found in other primates?
Two of the most important evolutionary principles accounting for the characteristics of animals are (1) common descent, and (2) adaptation driven by natural selection. If we are all related to one another, and to all other species, by virtue of common descent, then one might expect to find similarities between humans and their closest primate relatives. This phylogenetic approach has a long history in psychology: it prompts the search for phylogenetic continuities implied by the inheritance of homologous features from common ancestors.
An adaptationist approach to psychology leads to the search for adaptive design, which usually entails the examination of niche-differentiated mental abilities unique to the species being investigated. George Williams's 1966 book, Adaptation and Natural Selection, clarified the logic of adaptationism. In so doing, this work laid the foundations of modern evolutionary psychology. Evolutionary psychology can be thought of as the application of adaptationist logic to the study of the architecture of the human mind.
Why does structure reflect function? In evolutionary biology, there are several different levels of explanation that are complementary and mutually compatible. Explanation at one level (e.g., adaptive function) does not preclude or invalidate explanations at another (e.g., neural, cognitive, social, cultural, economic). EPs use theories of adaptive function to guide their investigations of phenotypic structures. Why is this possible?
The evolutionary process has two components: chance and natural selection. Natural selection is the only component of the evolutionary process that can introduce complex functional organization in to a species' phenotype (Dawkins, 1986; Williams, 1966).
The function of the brain is to generate behavior that is sensitively contingent upon information from an organism's environment. It is, therefore, an information-processing device. Neuroscientists study the physical structure of such devices, and cognitive psychologists study the information-processing programs realized by that structure. There is, however, another level of explanation -- a functional level. In evolved systems, form follows function. The physical structure is there because it embodies a set of programs; the programs are there because they solved a particular problem in the past. This functional level of explanation is essential for understanding how natural selection designs organisms.
An organism's phenotypic structure can be thought of as a collection of "design features" -- micro-machines, such as the functional components of the eye or liver. Over evolutionary time, new design features are added or discarded from the species' design because of their consequences. A design feature will cause its own spread over generations if it has the consequence of solving adaptive problems: cross-generationally recurrent problems whose solution promotes reproduction, such as detecting predators or detoxifying poisons. If a more sensitive retina, which appeared in one or a few individuals by chance mutation, allows predators to be detected more quickly, individuals who have the more sensitive retina will produce offspring at a higher rate than those who lack it. By promoting the reproduction of its bearers, the more sensitive retina thereby promotes its own spread over the generations, until it eventually replaces the earlier-model retina and becomes a universal feature of that species' design.
Hence natural selection is a feedback process that "chooses" among alternative designs on the basis of how well they function. It is a hill-climbing process, in which a design feature that solves an adaptive problem well can be outcompeted by a new design feature that solves it better. This process has produced exquisitely engineered biological machines -- the vertebrate eye, photosynthetic pigments, efficient foraging algorithms, color constancy systems -- whose performance is unrivaled by any machine yet designed by humans.
By selecting designs on the basis of how well they solve adaptive problems, this process engineers a tight fit between the function of a device and its structure. To understand this causal relationship, biologists had to develop a theoretical vocabulary that distinguishes between structure and function. In evolutionary biology, explanations that appeal to the structure of a device are sometimes called "proximate" explanations. When applied to psychology, these would include explanations that focus on genetic, biochemical, physiological, developmental, cognitive, social, and all other immediate causes of behavior. Explanations that appeal to the adaptive function of a device are sometimes called "distal" or "ultimate" explanations, because they refer to causes that operated over evolutionary time.
Knowledge of adaptive function is necessary for carving nature at the joints. An organism's phenotype can be partitioned into adaptations, which are present because they were selected for, by-products, which are present because they are causally coupled to traits that were selected for (e.g., the whiteness of bone), and noise, which was injected by the stochastic components of evolution. Like other machines, only narrowly defined aspects of organisms fit together into functional systems: most ways of describing the system will not capture its functional properties. Unfortunately, some have misrepresented the well-supported claim that selection creates functional organization as the obviously false claim that all traits of organisms are funtional -- something no sensible evolutionary biologist would ever maintain. Furthermore, not all behavior engaged in by organisms is adaptive. A taste for sweet may have been adaptive in ancestral environments where vitamin-rich fruit was scarce, but it can generate maladaptive behavior in a modern environment flush with fast-food restaurants. Moreover, once an information-processing mechanism exists, it can be deployed in activities that are unrelated to its original function -- because we have evolved learning mechanisms that cause language acquisition, we can learn to write. But these learning mechanisms were not selected for because they caused writing.
Design evidence. Adaptations are problem-solving machines, and can be identified using the same standards of evidence that one would use to recognize a human-made machine: design evidence. One can identify a machine as a TV rather than a stove by finding evidence of complex functional design: showing, e.g., that it has many coordinated design features (antennaes, cathode ray tubes, etc.) that are complexly specialized for transducing TV waves and transforming them into a color bit map (a configuration that is unlikely to have risen by chance alone), whereas it has virtually no design features that would make it good at cooking food. Complex functional design is the hallmark of adaptive machines as well. One can identify an aspect of the phenotype as an adaptation by showing that (1) it has many design features that are complexly specialized for solving an adaptive problem, (2) these phenotypic properties are unlikely to have arisen by chance alone, and (3) they are not better explained as the by-product of mechanisms designed to solve some alternative adaptive problem. Finding that an architectural element solves an adaptive problem with "reliability, efficiency, and economy" is prima facie evidence that one has located an adaptation (Williams, 1966).
Design evidence is important not only for explaining why a known mechanism exists, but also for discovering new mechanisms, ones that no one had thought to look for. EPs also use theories of adaptive function heuristically, to guide their investigations of phenotypic design.
Those who study species from an adaptationist perspective adopt the stance of an engineer. In discussing sonar in bats, e.g., Dawkins proceeds as follows: "...I shall begin by posing a problem that the living machine faces, then I shall consider possible solutions to the problem that a sensible engineer might consider; I shall finally come to the solution that nature has actually adopted" (1986, pp. 21-22). Engineers figure out what problems they want to solve, and then design machines that are capable of solving these problems in an efficient manner. Evolutionary biologists figure out what adaptive problems a given species encountered during its evolutionary history, and then ask themselves, "What would a machine capable of solving these problems well under ancestral conditions look like?" Against this background, they empically explore the design features of the evolved machines that, taken together, comprise an organism. Definitions of adaptive problems do not, of course, uniquely specify the design of the mechanisms that solve them. Because there are often multiple ways of acheiving any solution, empirical studies are needed to decide "which nature has actually adopted". But the more precisely one can define an adaptive information-processing problem -- the "goal" of processing -- the more clearly one can see what a mechanism capable of producing that solution would have to look like. This research strategy has dominated the study of vision, for example, so that it is now commonplace to think of the visual system as a collection of functionally integrated computational devices, each specialized for solving a different problem in scene analysis -- judging depth, detecting motion, analyzing shape from shading, and so on. In our own research, we have applied this strategy to the study of social reasoning (see below).
To fully understand the concept of design evidence, we need to consider how an adaptationist thinks about nature and nurture.
Nature and nurture: An adaptationist perspective
Debates about the "relative contribution" during development of "nature" and "nurture" have been among the most contentious in psychology. The premises that underlie these debates are flawed, yet they are so deeply entrenched that many people have difficulty seeing that there are other ways to think about these issues.
Evolutionary psychology is not just another swing of the nature/nurture pendulum. A defining characteristic of the field is the explicit rejection of the usual nature/nurture dichotomies -- instinct vs. reasoning, innate vs. learned, biological vs. cultural. What effect the environment will have on an organism depends critically on the details of its evolved cognitive architecture. For this reason, coherent "environmentalist" theories of human behavior all make "nativist" claims about the exact form of our evolved psychological mechanisms. For an EP, the real scientific issues concern the design, nature, and number of these evolved mechanisms, not "biology versus culture" or other malformed oppositions.
There are several different "nature-nurture" issues, which are usually conflated. Let's pull them apart and look at them separately, because some of them are non-issues whereas others are real issues.
Focus on architecture. At a certain level of abstraction, every species has a universal, species-typical evolved architecture. For example, one can open any page of the medical textbook, Gray's Anatomy, and find the design of this evolved architecture described down to the minutest detail -- not only do we all have a heart, two lungs, a stomach, intestines, and so on, but the book will describe human anatomy down to the particulars of nerve connections. This is not to say there is no biochemical individuality: No two stomachs are exactly alike -- they vary a bit in quantitative properties, such as size, shape, and how much HCl they produce. But all humans have stomachs and they all have the same basic functional design -- each is attached at one end to an esophagus and at the other to the small intestine, each secretes the same chemicals necessary for digestion, and so on. Presumably, the same is true of the brain and, hence, of the evolved architecture of our cognitive programs -- of the information-processing mechanisms that generate behavior. Evolutionary psychology seeks to characterize the universal, species-typical architecture of these mechanisms.
The cognitive architecture, like all aspects of the phenotype from molars to memory circuits, is the joint product of genes and environment. But the development of architecture is buffered against both genetic and environmental insults, such that it reliably develops across the (ancestrally) normal range of human environments. EPs do not assume that genes play a more important role in development than the environment does, or that "innate factors" are more important than "learning". Instead, EPs reject these dichotomies as ill-conceived.
Evolutionary psychology is not behavior genetics. Behavior geneticists are interested in the extent to which differences between people in a given environment can be accounted for by differences in their genes. EPs are interested in individual differences only insofar as these are the manifestation of an underlying architecture shared by all human beings. Because their genetic basis is universal and species-typical, the heritability of complex adaptations (of the eye, for example) is usually low, not high. Moreover, sexual recombination constrains the design of genetic systems, such that the genetic basis of any complex adaptation (such as a cognitive mechanism) must be universal and species-typical (Tooby and Cosmides, 1990b). This means the genetic basis for the human cognitive architecture is universal, creating what is sometimes called the psychic unity of humankind. The genetic shuffle of meiosis and sexual recombination can cause individuals to differ slightly in quantitative properties that do not disrupt the functioning of complex adaptations. But two individuals do not differ in personality or morphology because one has the genetic basis for a complex adaptation that the other lacks. The same principle applies to human populations: from this perspective, there is no such thing as "race".
In fact, evolutionary psychology and behavior genetics are animated by two radically different questions:
What is the universal, evolved architecture that we all share by virtue of being humans? (evolutionary psychology)
Given a large population of people in a specific environment, to what extent can differences between these people be accounted for by differences in their genes? (behavior genetics)
The second question is usually answered by computing a heritability coefficient, based on (for example) studies of identical and fraternal twins. "Which contributes more to nearsightedness, genes or environment" (an instance of the second question), has no fixed answer: the "heritability" of a trait can vary from one place to the next, precisely because environments do affect development.
A heritability coefficient measures sources of variance in a population (for example, in a forest of oaks, to what extent are differences in height correlated with differences in sunlight, all else equal?). It tells you nothing about what caused the development of an individual. Let's say that for height, 80% of the variance in a forest of oaks is caused by variation in their genes. This does not mean that the height of the oak tree in your yard is "80% genetic". (What could this possibly mean? Did genes contribute more to your oak's height than sunlight? What percent of its height was caused by nitrogen in the soil? By rainfall? By the partial pressure of CO2?) When applied to an individual, such percents are meaningless, because all of these factors are necessary for a tree to grow. Remove any one, and the height will be zero.
Joint product of genes and environment. Confusing individuals with populations has led many people to define "the" nature-nurture question in the following way: What is more important in determining an (individual) organism's phenotype, its genes or its environment?
Any developmental biologist knows that this is a meaningless question. Every aspect of an organism's phenotype is the joint product of its genes and its environment. To ask which is more important is like asking, Which is more important in determining the area of a rectangle, the length or the width? Which is more important in causing a car to run, the engine or the gasoline? Genes allow the environment to influence the development of phenotypes.
Indeed, the developmental mechanisms of many organisms were designed by natural selection to produce different phenotypes in different environments. Certain fish can change sex, for example. Blue-headed wrasse live in social groups consisting of one male and many females. If the male dies, the largest female turns into a male. The wrasse are designed to change sex in response to a social cue -- the presence or absence of a male.
With a causal map of a species' developmental mechanisms, you can change the phenotype that develops by changing its environment. Imagine planting one seed from an arrowleaf plant in water, and a genetically identical seed on dry land. The one in water would develop wide leaves, and the one on land would develop narrow leaves. Responding to this dimension of environmental variation is part of the species' evolved design. But this doesn't mean that just any aspect of the environment can affect the leaf width of an arrowleaf plant. Reading poetry to it doesn't affect its leaf width. By the same token, it doesn't mean that it is easy to get the leaves to grow into just any shape: short of a pair of scissors, it is probably very difficult to get the leaves to grow into the shape of the Starship Enterprise.
People tend to get mystical about genes; to treat them as "essences" that inevitably give rise to behaviors, regardless of the environment in which they are expressed. But genes are simply regulatory elements, molecules that arrange their surrounding environment into an organism. There is nothing magical about the process: DNA is transcribed into RNA; within cells, at the ribosomes, the RNA is translated into proteins -- the enzymes -- that regulate development. There is no aspect of the phenotype that cannot be influenced by some environmental manipulation. It just depends on how ingenious or invasive you want to be. If you drop a human zygote (a fertilized human egg) into liquid nitrogen, it will not develop into an infant. If you were to shoot electrons at the zygote's ribosomes in just the right way, you could influence the way in which the RNA is translated into proteins. By continuing to do this you could, in principle, cause a human zygote to develop into a watermelon or a whale. There is no magic here, only causality.
Present at birth? Sometimes people think that to show that an aspect of the phenotype is part of our evolved architecture, one must show that it is present from birth. But this is to confuse an organism's "initial state" with its evolved architecture. Infants do not have teeth at birth -- they develop them quite awhile after birth. But does this mean they "learn" to have teeth? What about breasts? Beards? One expects organisms to have mechanisms that are adapted to their particular life stage (consider the sea squirt!) -- after all, the adaptive problems an infant faces are different from those an adolescent faces.
This misconception frequently leads to misguided arguments. For example, people think that if they can show that there is information in the culture that mirrors how people behave, then that is the cause of their behavior. So if they see that men on TV have trouble crying, they assume that their example is causing boys to be afraid to cry. But which is cause and which effect? Does the fact that men don't cry much on TV teach boys to not cry, or does it merely reflect the way boys normally develop? In the absence of research on the particular topic, there is no way of knowing. (To see this, just think about how easy it would be to argue that girls learn to have breasts. Consider the peer pressure during adolescence for having breasts! the examples on TV of glamorous models! -- the whole culture reinforces the idea that women should have breasts, therefore...adolescent girls learn to grow breasts.)
In fact, an aspect of our evolved architecture can, in principle, mature at any point in the life-cycle, and this applies to the cognitive programs of our brain just as much as it does to other aspects of our phenotype.
Is domain-specificity politically incorrect? Sometimes people favor the notion that everything is "learned" -- by which they mean "learned via general purpose circuits" -- because they think it supports democratic and egalitarian ideals. They think it means anyone can be anything. But the notion that anyone can be anything gets equal support, whether our circuits are specialized or general. When we are talking about a species' evolved architecture, we are talking about something that is universal and species-typical -- something all of us have. This is why the issue of specialization has nothing to do with "democratic, egalitarian ideals" -- we all have the same basic biological endowment, whether it is in the form of general purpose mechanisms or special purpose ones. If we all have a special purpose "language acquisition device", for example (see Pinker, this volume), we are all on an "equal footing" when it comes to learning language, just as we would be if we learned language via general purpose circuits.
"Innate" is not the opposite of "learned". For EPs, the issue is never "learning" versus "innateness" or "learning" versus "instinct". The brain must have a certain kind of structure for you to learn anything at all -- after all, three pound bowls of oatmeal don't learn, but three pound brains do. If you think like an engineer, this will be clear. To learn, there must be some mechanism that causes this to occur. Since learning cannot occur in the absence of a mechanism that causes it, the mechanism that causes it must itself be unlearned -- must be "innate". Certain learning mechanisms must therefore be aspects of our evolved architecture that reliably develop across the kinds of environmental variations that humans normally encountered during their evolutionary history. We must, in a sense, have what you can think of as "innate learning mechanisms" or "learning instincts". The interesting question is what are these unlearned programs? Are they specialized for learning a particular kind of thing, or are they designed to solve more general problems? This brings us back to Principle 4.
Specialized or general purpose? One of the few genuine nature-nurture issues concerns the extent to which a mechanism is specialized for producing a given outcome. Most nature/nurture dichotomies disappear when one understands more about developmental biology, but this one does not. For EPs, the important question is, What is the nature of our universal, species-typical evolved cognitive programs? What kind of circuits do we actually have?
The debate about language acquisition brings this issue into sharp focus: Do general purpose cognitive programs cause children to learn language, or is language learning caused by programs that are specialized for performing this task? This cannot be answered a priori. It is an empirical question, and the data collected so far suggest the latter (Pinker, 1994, this volume).
For any given behavior you observe, there are three possibilities:
It is the product of general purpose programs (if such exist);
It is the product of cognitive programs that are specialized for producing that behavior; or
It is a by-product of specialized cognitive programs that evolved to solve a different problem. (Writing, which is a recent cultural invention, is an example of the latter.)
More nature allows more nurture. There is not a zero-sum relationship between "nature" and "nurture". For EPs, "learning" is not an explanation -- it is a phenomenon that requires explanation. Learning is caused by cognitive mechanisms, and to understand how it occurs, one needs to know the computational structure of the mechanisms that cause it. The richer the architecture of these mechanisms, the more an organism will be capable of learning -- toddlers can learn English while (large-brained) elephants and the family dog cannot because the cognitive architecture of humans contains mechanisms that are not present in that of elephants or dogs. Furthermore, "learning" is a unitary phenomenon: the mechanisms that cause the acquisition of grammar, for example, are different from those that cause the acquisition of snake phobias. (The same goes for "reasoning".)
What evolutionary psychology is not. For all the reasons discussed above, EPs expect the human mind will be found to contain a large number of information-processing devices that are domain-specific and functionally specialized. The proposed domain-specificity of many of these devices separates evolutionary psychology from those approaches to psychology that assume the mind is composed of a small number of domain general, content-independent, "general purpose" mechanisms -- the Standard Social Science Model.
It also separates evolutionary psychology from those approaches to human behavioral evolution in which it is assumed (usually implicitly) that "fitness-maximization" is a mentally (though not consciously) represented goal, and that the mind is composed of domain general mechanisms that can "figure out" what counts as fitness-maximizing behavior in any environment -- even evolutionarily novel ones (Cosmides and Tooby, 1987; Symons, 1987, 1992). Most EPs acknowledge the multipurpose flexibility of human thought and action, but believe this is caused by a cognitive achitecture that contains a large number of evolved "expert systems".
Reasoning instincts: An example
In some of our own research, we have been exploring the hypothesis that the human cognitive architecture contains circuits specialized for reasoning about adaptive problems posed by the social world of our ancestors. In categorizing social interactions, there are two basic consequences humans can have on each other: helping or hurting, bestowing benefits or inflicting costs. Some social behavior is unconditional: one nurses an infant without asking it for a favor in return, for example. But most social acts are conditionally delivered. This creates a selection pressure for cognitive designs that can detect and understand social conditionals reliably, precisely, and economcally (Cosmides, 1985, 1989; Cosmides & Tooby, 1989, 1992). Two major categories of social conditionals are social exchange and threat -- conditional helping and conditional hurting -- carried out by individuals or groups on individuals or groups. We initially focused on social exchange (for review, see Cosmides & Tooby, 1992).
We selected this topic for several reasons:
Many aspects of the evolutionary theory of social exchange (sometimes called cooperation, reciprocal altruism, or reciprocation) are relatively well-developed and unambiguous. Consequently, certain features of the functional logic of social exchange could be confidently relied on in constructing hypotheses about the structure of the information-processing procedures that this activity requires.
Complex adaptations are constructed in response to evolutionarily long-enduring problems. Situations involving social exchange have constituted a long-enduring selection pressure on the hominid line: evidence from primatology and paleoanthropology suggests that our ancestors have engaged in social exchange for at least several million years.
Social exchange appears to be an ancient, pervasive and central part of human social life. The universality of a behavioral phenotype is not a sufficient condition for claiming that it was produced by a cognitive adaptation, but it is suggestive. As a behavioral phenotype, social exchange is as ubiquitous as the human heartbeat. The heartbeat is universal because the organ that generates it is everywhere the same. This is a parsimonious explanation for the unversality of social exchange as well: the cognitive phenotype of the organ that generates it is everywhere the same. Like the heart, its development does not seem to require environmental conditions (social or otherwise) that are idiosyncratic or culturally contingent.
Theories about reasoning and rationality have played a central role in both cognitive science and the social sciences. Research in this area can, as a result, serve as a powerful test of the central assumption of the Standard Social Science Model: that the evolved architecture of the mind consists solely or predominantly of a small number of content-independent, general-purpose mechanisms.
The evolutionary analysis of social exchange parallels the economist's concept of trade. Sometimes known as "reciprocal altruism", social exchange is an "I'll scratch your back if you scratch mine" principle. Economists and evolutionary biologists had already explored constraints on the emergence or evolution of social exchange using game theory, modeling it as a repeated Prisoners' Dilemma. One important conclusion was that social exchange cannot evolve in a species or be stably sustained in a social group unless the cognitive machinery of the participants allows a potential cooperator to detect individuals who cheat, so that they can be excluded from future interactions in which they would exploit cooperators (e.g., Axelrod, 1984; Axelrod & Hamilton, 1981; Boyd, 1988; Trivers, 1971; Williams, 1966). In this context, a cheater is an individual who accepts a benefit without satisfying the requirements that provision of that benefit was made contingent upon.
Such analyses provided a principled basis for generating detailed hypotheses about reasoning procedures that, because of their domain-specialized structure, would be well-designed for detecting social conditionals, interpreting their meaning, and successfully solving the inference problems they pose. In the case of social exchange, for example, they led us to hypothesize that the evolved architecture of the human mind would include inference procedures that are specialized for detecting cheaters.
To test this hypothesis, we used an experimental paradigm called the Wason selection task (Wason, 1966; Wason & Johnson-Laird, 1972). For about 20 years, psychologists had been using this paradigm (which was originally developed as a test of logical reasoning) to probe the structure of human reasoning mechanisms. In this task, the subject is asked to look for violations of a conditional rule of the form If P then Q. Consider the Wason selection task presented in Figure 3.
Figure 3.
Part of your new job for the City of Cambridge is to study the demographics of transportation. You read a previously done report on the habits of Cambridge residents that says: "If a person goes into Boston, then that person takes the subway."
The cards below have information about four Cambridge residents. Each card represents one person. One side of a card tells where a person went, and the other side of the card tells how that person got there. Indicate only those card(s) you definitely need to turn over to see if any of these people violate this rule.
|
|
|
|
From a logical point of view, the rule has been violated whenever someone goes to Boston without taking the subway. Hence the logically correct answer is to turn over the Boston card (to see if this person took the subway) and the cab card (to see if the person taking the cab went to Boston). More generally, for a rule of the form If P then Q, one should turn over the cards that represent the values P and not-Q (to see why, consult Figure 2).
If the human mind develops reasoning procedures specialized for detecting logical violations of conditional rules, this would be intuitively obvious. But it is not. In general, fewer than 25% of subjects spontaneously make this response. Moreover, even formal training in logical reasoning does little to boost performance on descriptive rules of this kind (e.g., Cheng, Holyoak, Nisbett & Oliver, 1986; Wason & Johnson-Laird, 1972). Indeed, a large literature exists that shows that people are not very good at detecting logical violations of if-then rules in Wason selection tasks, even when these rules deal with familiar content drawn from everyday life (e.g., Manktelow & Evans, 1979; Wason, 1983).
The Wason selection task provided an ideal tool for testing hypotheses about reasoning specializations designed to operate on social conditionals, such as social exchanges, threats, permissions, obligations, and so on, because (1) it tests reasoning about conditional rules, (2) the task structure remains constant while the content of the rule is changed, (3) content effects are easily elicited, and (4) there was already a body of existing experimental results against which performance on new content domains could be compared.
For example, to show that people who ordinarily cannot detect violations of conditional rules can do so when that violation represents cheating on a social contract would constitute initial support for the view that people have cognitive adaptations specialized for detecting cheaters in situations of social exchange. To find that violations of conditional rules are spontaneously detected when they represent bluffing on a threat would, for similar reasons, support the view that people have reasoning procedures specialized for analyzing threats. Our general research plan has been to use subjects' inability to spontaneously detect violations of conditionals expressing a wide variety of contents as a comparative baseline against which to detect the presence of performance-boosting reasoning specializations. By seeing what content-manipulations switch on or off high performance, the boundaries of the domains within which reasoning specializations successfully operate can be mapped.
The results of these investigations were striking. People who ordinarily cannot detect violations of if-then rules can do so easily and accurately when that violation represents cheating in a situation of social exchange (Cosmides, 1985, 1989; Cosmides & Tooby, 1989; 1992). This is a situation in which one is entitled to a benefit only if one has fulfilled a requirement (e.g., "If you are to eat those cookies, then you must first fix your bed"; "If a man eats cassava root, then he must have a tattoo on his chest"; or, more generally, "If you take benefit B, then you must satisfy requirement R"). Cheating is accepting the benefit specified without satisfying the condition that provision of that benefit was made contingent upon (e.g., eating the cookies without having first fixed your bed).
When asked to look for violations of social contracts of this kind, the adaptively correct answer is immediately obvious to almost all subjects, who commonly experience a "pop out" effect. No formal training is needed. Whenever the content of a problem asks subjects to look for cheaters in a social exchange -- even when the situation described is culturally unfamiliar and even bizarre -- subjects experience the problem as simple to solve, and their performance jumps dramatically. In general, 65-80% of subjects get it right, the highest performance ever found for a task of this kind. They choose the "benefit accepted" card (e.g., "ate cassava root") and the "cost not paid" card (e.g., "no tattoo"), for any social conditional that can be interpreted as a social contract, and in which looking for violations can be interpreted as looking for cheaters.
From a domain-general, formal view, investigating men eating cassava root and men without tattoos is logically equivalent to investigating people going to Boston and people taking cabs. But everywhere it has been tested (adults in the US, UK, Germany, Italy, France, Hong-Kong; schoolchildren in Ecuador, Shiwiar hunter-horticulturalists in the Ecuadorian Amazon), people do not treat social exchange problems as equivalent to other kinds of reasoning problems. Their minds distinguish social exchange contents, and reason as if they were translating these situations into representational primitives such as "benefit", "cost", "obligation", "entitlement", "intentional", and "agent." Indeed, the relevant inference procedures are not activated unless the subject has represented the situation as one in which one is entitled to a benefit only if one has satisfied a requirement.
Moreover, the procedures activated by social contract rules do not behave as if they were designed to detect logical violations per se; instead, they prompt choices that track what would be useful for detecting cheaters, whether or not this happens to correspond to the logically correct selections. For example, by switching the order of requirement and benefit within the if-then structure of the rule, one can elicit responses that are functionally correct from the point of view of cheater detection, but logically incorrect (see Figure 4). Subjects choose the benefit accepted card and the cost not paid card -- the adaptively correct response if one is looking for cheaters -- no matter what logical category these cards correspond to.
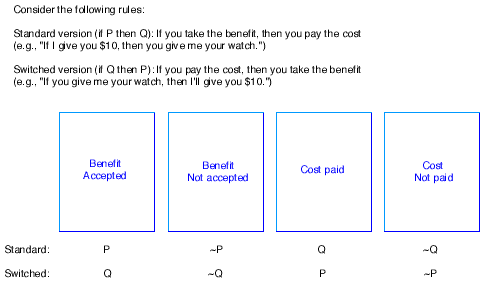
Figure 4: Generic Structure of a Social Contract.
To show that an aspect of the phenotype is an adaptation, one needs to demonstrate a fit between form and function: one needs design evidence. There are now a number of experiments comparing performance on Wason selection tasks in which the conditional rule either did or did not express a social contract. These experiments have provided evidence for a series of domain-specific effects predicted by our analysis of the adaptive problems that arise in social exchange. Social contracts activate content-dependent rules of inference that appear to be complexly specialized for processing information about this domain. Indeed, they include subroutines that are specialized for solving a particular problem within that domain: cheater detection. The programs involved do not operate so as to detect potential altruists (individuals who pay costs but do not take benefits), nor are they activated in social contract situations in which errors would correspond to innocent mistakes rather than intentional cheating. Nor are they designed to solve problems drawn from domains other than social exchange; for example, they will not allow one to detect bluffs and double crosses in situations of threat, nor will they allow one to detect when a safety rule has been violated. The pattern of results elicited by social exchange content is so distinctive that we believe reasoning in this domain is governed by computational units that are domain specific and functionally distinct: what we have called social contract algorithms (Cosmides, 1985, 1989; Cosmides & Tooby, 1992).
There is, in other words, design evidence. The programs that cause reasoning in this domain have many coordinated features that are complexly specialized in precisely the ways one would expect if they had been designed by a computer engineer to make inferences about social exchange reliably and efficiently: configurations that are unlikely to have arisen by chance alone. Some of these design features are listed in Table 1, as well as a number of by-product hypotheses that have been empirically eliminated. (For review, see Cosmides & Tooby, 1992; also Cosmides, 1985, 1989; Cosmides & Tooby, 1989; Fiddick, Cosmides, & Tooby, 1995; Gigerenzer & Hug, 1992; Maljkovic, 1987; Platt & Griggs, 1993.)
It may seem strange to study reasoning about a topic as emotionally charged as cheating -- after all, many people (starting with Plato) talk about emotions as if they were goo that clogs the gearwheels of reasoning EPs can address such topics, however, because most of them see no split between "emotion" and "cognition". There are probably many ways of conceptualizing emotions from an adaptationist point of view, many of which would lead to interesting competing hypotheses. One that we find useful is as follows: an emotion is a mode of operation of the entire cognitive system, caused by programs that structure interactions among different mechanisms so that they function particularly harmoniously when confronting cross-generationally recurrent situations -- especially ones in which adaptive errors are so costly that you have to respond appropriately the first time you encounter them (see Tooby & Cosmides, 1990a).
Their focus on adaptive problems that arose in our evolutionary past has led EPs to apply the concepts and methods of the cognitive sciences to many nontraditional topics: the cognitive processes that govern cooperation, sexual attraction, jealousy, parental love, the food aversions and timing of pregnancy sickness, the aesthetic preferences that govern our appreciation of the natural environment, coalitional aggression, incest avoidance, disgust, foraging, and so on (for review, see Barkow, Cosmides, & Tooby, 1992). By illuminating the programs that give rise to our natural competences, this research cuts straight to the heart of human nature.
Acknowledgements:
We would like to thank Martin Daly, Irv DeVore, Steve Pinker, Roger Shepard, Don Symons, and Margo Wilson for many fruitful discussions of these issues, and William Allman for suggesting the phrase, "Our modern skulls house a stone age mind", which is a very apt summary of our position. We are grateful to the James S. McDonnell Foundation and NSF Grant BNS9157-499 to John Tooby, for their financial support during the preparation of this chapter.
Copyright John Tooby and Leda Cosmides, 1997. Original Paper at
UCSB Updated January 13, 2023
^ Top ^
|
|
|
